01:07 Антигены I класса главного комплекса гистосовместимости | |
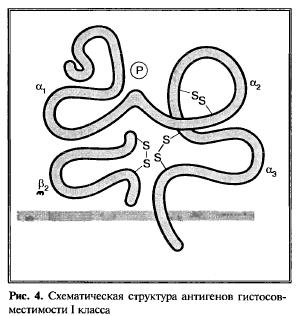
Антигены I класса ГКГ в норме экспрессируются практически всеми ядросодержащими клетками (исключение составляют клетки ранних стадий эмбрионального развития). Антигены ГКГ представляют собой универсальные структуры, количество которых колеблется в зависимости от вида ткани и достигает максимума на мембране лимфоцитов всех лимфоидных тканей (лимфатических узлов, селезенки), а также в периферической крови. Значительно ниже уровень экспрессии антигенов I класса ГКГ в клетках печени, почек и эндокринных органов. Особенности тканей и их функциональное состояние, возможность развития той или иной патологии также влияют на уровень экспрессии антигенов I класса ГКГ. Клетки, лишенные антигенов ГКГ, считаются мутантными. Беспрецедентный полиморфизм антигенов ГКГ внутри вида обеспечивает уникальность и неповторимость антигенной структуры отдельных индивидуумов одного и того же вида; контроль за этим полиморфизмом осуществляют гены ГКГ. Необходимо также учитывать, что исходно в нормальных тканях уровень экспрессии антигенов ГКГ I класса различен и зависит от локализации и особенностей тех или иных клеток. Например, на клетках эпителия кишечника, гортани, молочной железы, легких уровень экспрессии антигенов I класса ГКГ обычно высокий, на клетках скелетных мышц и слизистой желудка — невысокий, а на клетках центральной нервной системы эти антигены практически не выявляются. Гетерогенность клеточного состава тех или иных органов или тканей, в свою очередь, определяет и возможные различия в экспрессии антигенов I класса ГКГ различными клетками. Важную роль в этом играют особенности микроокружения, в частности продукция цитокинов, которые по-разному влияют на экспрессию антигенов I класса ГКГ. Молекулы антигенов I класса ГКГ представлены различными локусами: А, В, С — классические молекулы с выраженным полиморфизмом, а также локусы G, E и F, известные как неклассические молекулы антигенов I класса ГКГ; к неклассическим молекулам относятся и CD1d. И классические молекулы антигенов ГКГ I класса, и неклассические антигены локуса G могут находиться в растворимой форме — sHLA-A, sHLA-B, sHLA-C, а также sHLA-G. Основные структурные особенности антигенов I класса ГКГ таковы. Молекула антигенов этого класса представляет собой интегральный мембранный гликопротеин (гетеродимер с молекулярной массой 45 кД) и состоит из тяжелой α-цепи, в состав которой входят α1-, α2- и α3-домены. Домены α1 и α2 могут непосредственно связываться с опухолевыми пептидами, в то время как а3-домен содержит неполиморфный регион — лиганд для цитотоксических T-клеток, который взаимодействует с рецептором СD8+-лимфоцитв в и гомологичен контактному участку Ig. Функционирование молекул I класса ГКГ во многом связано с β2-микроглобулином (β2m) который играет важную роль в особенностях α-цепи и представляет собой растворимую легкую цепь. В литературе все чаще появляются сообщения, авторы которых пытаются найти связь между экспрессией антигенов I класса ГКГ и геном β2m. Полученные данные разноречивы. Тем не менее постановка этого вопроса имеет серьезное обоснование, базирующееся на таких двух убедительных фактах. Первый — независимо от того, можно ли в настоящее время утверждать наличие связи между снижением экспрессии ГКГ и β2m, показано, что опухолевые пептиды могут непосредственно связываться с β2m, образуя комплекс с тяжелой цепью молекулы антигена ГКГ I класса. В частности, такой способностью обладает HLA-A2-peстриктированный эпитоп, связывающийся с N-концами β2m, который затем распознается ЦТЛ. Второй — аномалии в экспрессии β2m часто сочетаются с резистентностью к действию ЦТЛ. Что касается выявления каких-либо корреляции между экспрессией антигенов I класса ГКГ и β2m, то, как уже указывалось, эти данные неоднородны. Исследование большого числа различных опухолей (меланома, рак кишечника, носоглотки и др.) показало, что в подавляющем количестве наблюдений экспрессия антигенов I класса ГКГ уменьшалась. В одних случаях это сочеталось с мутацией гена β2m, а в других — нет. Из этого следует, что авторы приведенных данных не рассматривают соматические мутации гена β2m как главный механизм снижения уровня антигенов I класса ГКГ. В противоположность такой точке зрения при изучении экспрессии антигенов I класса ГКГ (А, В, С) параллельно с геном β2m другие авторы показали, что уменьшение экспрессии указанных антигенов при первичных карциномах рака молочной железы в 40 % случаев сочеталось и со снижением экспрессии гена β2m по сравнению с этим показателем для нормальных тканей. Лишь в 12 % появление β2m было сравнимо с нормой; снижение экспрессии β2m не сопровождалось дефектами гена β2m. Исследование молекулярных механизмов уменьшения экспрессии антигенов I класса ГКГ дало основание для заключения, что такое снижение представляет собой феномен, который происходит главным образом на посттранскрипционном уровне и может влиять на экспрессию гена β2m. Более однозначную трактовку наличия β2m высказывают другие авторы. Так, показано, что в значительном количестве клеток различных линий злокачественных опухолей, включая меланому, рак почки и другие, резко снижен уровень экспрессии антигенов I класса ГКГ и параллельно экспрессия β2m либо ослаблена, либо этот микроглобулин вообще не экспрессируется. Наконец, нельзя не отметить и данные, согласно которым отсутствие экспрессии или невысокий уровень β2m у мышей некоторых линий сочетается с дефектом созревания CD4 CDS T-лимфоцитов, экспрессирующих αβTCR и дефектом цитотоксичности Т-лимфоцитов. Из этих данных следует, что экспрессия молекул I класса ГКГ играет ключевую роль в положительной селекции Т-клеток, в частности тех из них, которые в период тимического созревания экспрессируют α- и β-цепи TCR. Несмотря на указанную противоречивость данных, изучение β2m, его исследование при карциноме носоглотки показало достоверное повышение уровня этого белка с различиями на отдельных этапах процесса, при распространении опухоли и метастазах. Повышение уровня β2m наиболее часто наблюдалось при низкодифференцированных формах этой опухоли, однако, по мнению авторов исследований, диагностическая значимость этого маркера низкая. Приведенные данные, несмотря на их некоторую разноречивость, свидетельствуют о том, что экспрессия β2m в злокачественно трансформированных клетках в значительном количестве случаев ассоциируется с дефектами распознавания и снижением цитотоксичности, что объясняет заслуженный интерес к изучению роли β2m в процессе распознавания опухолевых антигенов. Весьма вероятно, что дальнейшие исследования в этом направлении могут послужить базой не только для прогнозирования течения опухолевого процесса, но и для подходов к регуляции индукции иммунологического ответа. Схематически структура классических антигенов I класса ГКГ представлена на рис. 4.
Стремление к выяснению механизмов снижения экспрессии антигенов I класса ГКГ не ограничивается поисками связи с мутациями гена p2m. В частности, показано, что это может быть обусловлено потерей гетерозиготности (loss of heterozigosity — LOH) на 6р21 хромосоме. Этот механизм приводит к необратимому снижению уровня HLA-гаплотипа в различных опухолях и, несмотря на недостаточную изученность, может быть серьезным препятствием для терапевтического эффекта при иммунизации опухолевыми антигенами. Уменьшение гетерозиготности было обнаружено в образцах, полученных из опухолей носоглотки, кишечника, меланомы, что позволило авторам на основании большого исследованного материала разделить опухоли на LOH-нега-тивные и LOH-позитивные для выявления больных, которых можно рассматривать как перспективных для иммунотерапии. Для реализации процесса распознавания антигены I класса ГКГ в комплексе с опухолевыми пептидами должны быть доставлены на поверхность опухолевой клетки. Транспорт этого комплекса, как правило, может быть осуществлен только при наличии белков-транспортеров — ТАР (transporter antigene proteines). ТАР представляет собой гетеродимер, относящийся к субсемейству трансмембранных транспортеров, синтезируется в цитозоле, где связан с комплексом, включающим и α-цепи ГКГ, опухолевого пептида, β2m, и транспортирует этот комплекс в эндоплазматический ретикулум, где и происходит процессинг. В настоящее время известны две субъединицы этого белка — TAP-1 и ТАР-2. Значение TAP-1 и ТАР-2 в процессе распознавания не ограничивается транспортом указанного комплекса, так как наряду с этим они обеспечивают и организацию молекул ГКГ. Регуляция активности белков-транспортеров осуществляется факторами PSF1 и PSF2 (peptide suppy factors). Молекула ГКГ I класса взаимодействует с белком-транспортером благодаря молекуле, известной как тапазин, которая кодируется геном, связанным с ГКГ. Экспрессия тапазина в ряде случаев может корректировать дефекты распознавания ЦТЛ, что свидетельствует о важной роли этого белка в HLA-1-рестриктированном распознавании. Образовавшийся в последующем указанный выше тример из цитозоля через аппарат Гольджи транспортируется на поверхность опухолевой клетки и презентирует соответствующие эпитопы рецепторам СD8+Т-лимфоцитов. Рис. 5 иллюстрирует участие транспортных белков в перемещении комплекса антигены ГКГ — антигены опухоли. В плане общих представлений о функционировании ТАР имеют значение также данные, полученные в последнее время при исследовании клеток меланомы. Из них следует, что появление точечных мутаций в генах, кодирующих антигены I класса ГКГ нарушает транспортную способность ТАР, что может препятствовать распознаванию ЦТЛ и рассматривается как еще одна причина ускользания опухоли из-под иммунологического контроля.
Эффективность презентации антигенов в комплексе с молекулами I класса ГКГ ЦТЛ зависит не только от наличия экспрессии ТАР, но и от их функциональной активности. Молекулярные механизмы нарушения функциональной активности ТАР изучены недостаточно. Однако в настоящее время уже есть сведения о некоторых механизмах нарушения экспрессии и функциональной активности ТАР. Предполагается, что такие нарушения могут быть обусловлены транслокацией и точечными мутациями в генах, кодирующих эти белки, что ведет к потере способности клетки презентировать антигены I класса ГКГ. Поэтому есть все основания полагать, что дефект этой системы можно считать одним из центральных в изменении экспрессии антигенов I класса ГКГ. Подтверждением этому служат результаты исследования клеток линии немелкоклеточной карциномы легкого, когда точечные мутации, сочетающиеся с нарушениями функции ТАР, были обнаружены в аденозин-трифосфатсвязывающем участке этого белка. He исключается также возможность наличия ингибиторов активности ТАР. Последнее предположение основано на том, что белок простого вируса герпеса ICP47 блокирует траспорт ТАР. В этой связи нельзя исключить и существование других ингибиторов активности ТАР как вирусного, так и другого происхождения. Следует обратить внимание также на неодинаковую степень значимости экспрессии ТАР в клетках высоко- и низкоиммуногенных опухолей. Так, изучение презентации пептидов вирусиндуцированных опухолей мышей линии С57В1/6 показало, что эффективность презентации пептида слабоиммуногенными опухолями четко зависит от экспрессии ТАР, в то время как выраженной зависимости от презентации пептидов высокоиммуногенными опухолями не прослеживается. Факт TAP-независимого распознавания нуждался в объяснении, возможность которого появилась лишь в самое последнее время благодаря работам Т. Fiedler и сотрудников. Им удалось получить данные, согласно которым в случаях дефекта ТАР презентация опухолевых антигенов с участием молекул CD1d остается неизмененной. В связи с этими данными авторы считают возможным рассматривать презентацию с участием CD1d как дополнительный механизм распознавания. Стали известны и молекулярные механизмы снижения функциональной активности ТАР человека и мышей, выявлены также структуры, которые обеспечивают активность этих белков-транспортеров. В частности, при изучении аминокислотной последовательности ТАР было установлено, что наличие глютаминовой кислоты в позиции 263 (Glu-263) обеспечивает их транспортную функцию. Снижение функциональной активности может быть также связано с нарушением стабильности гена мРНК, ответственного за презентацию антигена, что нередко сочетается и с уменьшением экспрессии антигенов I класса ГКГ. Изменение функциональной активности транспортных белков может приводить к нарушению процессинга антигенов. Об этом свидетельствует недавно установленный факт, полученный при исследовании карциномы почки; степень выраженности таких дефектов в клетках отдельных линий карциномы почки отличалась большой вариабельностью. Важно отметить, что частота обнаружения дефектов ТАР в различных опухолях неодинакова. Если они достаточно часто выявляются при меланомах, карциноме почки, то при раке легкого и карциномах кишечника снижение активности ТАР либо не наблюдалось, либо было слабо выраженным. Данные о неодинаковом уровне повреждений функциональной активности ТАР в различных опухолях представляются важными не только потому, что еще раз иллюстрируют биологические особенности опухолевых клеток, но и ориентируют на поиск механизмов, повреждение которых также может способствовать нарушению представления опухолевых антигенов. Важная роль экспрессии ТАР и должный уровень их функциональной активности для процесса распознавания опухолевых антигенов делает понятным, почему недостаточность этих транспортных белков очень существенно влияет на индукцию иммунологического ответа на данные антигены. Уже появились сведения о том, что снижение уровня экспрессии ТАР может быть использовано и для оценки клинических особенностей течения опухолевого процесса, в частности его прогноза. Такие данные, например, были получены при изучении клеток меланомы, когда было отмечено, что прогрессирующее течение меланомы и ускользание ее от распознавания ЦТЛ сочеталось со снижением уровня экспрессии ТАР. Параллельные исследования ТАР-1, ТАР-2, LMP-2, LMP-7, антигенов I класса ГКГ и β2m показали, что не только изменения ТАР-1, а, возможно, и ТАР-2, могут быть независимыми прогностическими маркерами при росте первичных меланом. Наряду с белками-транспортерами — важными компонентами распознавания большое значение имеет еще одна группа белков вирусного происхождения. Речь идет о белках вируса Эпштейна—Барр — LMP (large multifunctional protease), которые принадлежат к новому классу регуляторов и представляют собой субъединицу 20S протеосомы. В настоящее время известны несколько субъединиц этого белка — LMP-1, LMP-2А, LMP-2B, LMP-7, LMP-10 с различной молекулярной массой; идентифицированы 9 генов, кодирующих эти белки. Экспрессия белков LMP выявлена в различных опухолях: назофарингальной карциноме, раке желудка и других злокачественных опухолях эпителиального происхождения, лимфогранулематозе, лимфоме Беркигга и др. Имеются наблюдения, что LMP-2 чаще других белков этого семейства, например LMP-7, экспрессируются клетками как первичных опухолей, так и метастазов. Понимание роли LMP следует из особенностей тех процессов, в которых они участвуют. В этом плане достаточно изучены субъединицы LMP-2A и LMP-2B, которые имеют сходную молекулярную организацию. Белок LMP-2A связан с тирозиновыми киназами семейства src и является для них субстратом, а тирозинфосфорилирование LMP-2A индуцирует процесс адгезии к белку экстрацеллюлярного матрикса — ECM (extra cellular matrix). Наряду с перечисленными белками, участие которых обязательно практически во всех случаях распознавания, в этом процессе могут принимать участие и другие белки — MECL-1, РА28-α, РА28-β, тапазин и др., которые регулируются генами, сцепленными с генами, контролирующими презентацию антигена. Исходя из этого постулируется, что HLA-1-дефицитный фенотип опухоли, например меланомы, связан с уменьшением количества множества компонентов, среди которых прежде всего следует отметить ТАР, LMP, РА28-α или PA28-β, в то время как экспрессия других компонентов, таких, как калретикулин, ER60, белок дисульфидизомераза, калнексин либо вообще не изменена, либо снижена. Дефекты ТАР и LMP чаще наблюдаются в клетках метастазов, чем первичных опухолей, что может быть обусловлено большей генетической нестабильностью этих клеток. В результате создаются условия для селекции клона опухолевых клеток, способных ускользать от распознавания, рестриктированного молекулами I класса ГКГ. Исследование молекулярных механизмов процесса распознавания не ограничивается пониманием его сущности. Так, при изучении меланомы получены данные, согласно которым определение ТАР и LMP может иметь и клиническое значение. Результаты параллельного исследования LMP-2, LMP-7, ТАР-1, ТАР-2, антигенов I класса ГКГ и β2m в клетках меланомы различной плотности свидетельствуют о том, что: 1) экспрессия указанных маркеров не коррелировала с плотностью опухоли; 2) уменьшение количества LMP и ТАР во многих случаях сочеталось с ослаблением экспрессии молекул ГКГ; 3) снижение уровня экспрессии ТАР-1 и ТАР-2 коррелировало с наличием метастазов. Эти факты дали авторам основание для закпючения, что уровень экспрессии TAP-1 и ТАР-2 можно рассматривать как независимый прогностический признак. Еще одним примером неблагоприятного сочетания снижения уровня экспрессии молекул ГКГ, белков-транспортеров и опухолевых антигенов служат следующие данные. Оказалось, что уменьшение экспрессии антигена меланомы MART-1/Melan-A, ТАР и молекул ГКГ I класса в клетках больных меланомой приводило в последующем к летальному исходу; иммунотерапия была неэффективной. Это объясняет, почему в настоящее время предпринимаются попытки использования результатов определения экспрессии белков ТАР и LMP в клинике. Однако несмотря на бесспорную значимость белков ТАР и LMP в процессе распознавания, имеются наблюдения, которые иллюстрируют возможность исключений. Как неоднократно отмечаюсь, снижение экспрессии ТАР, как правило, связано с уменьшением экспрессии антигенов I класса ГКГ. Наряду с этим известны случаи, когда такой параллелизм отсутствует, что подтверждают результаты изучения клеток двух линий карциномы носоглотки человека. В клетках обеих линий уменьшалась экспрессия LMP-2, ТАР-1, ТАР-2, LMP-7, молекул аллелей HLA-B. В клетках одной из линий — HSC5, несмотря на выраженное снижение уровня ТАР отмечена экспрессия молекул HLA-A2, что свидетельствует о возможности транспортировки антигенов ГКГ без участия ТАР. Весьма вероятно, что такая возможность зависит от ряда еще не известных особенностей внутриклеточных процессов, происходящих в той или иной опухолевой клетке. Поэтому существование даже единичных случаев транспортировки комплексов опухолевых пептидов и молекул ГКГ при отсутствии ТАР ставит перед исследователями задачу выяснения, при каких условиях осуществляется распознавание. Таким образом, можно констатировать, что ТАР и LMP — необходимые компоненты эффективного процесса распознавания опухолевых антигенов. Снижение уровня экспрессии этих белков и их функциональной активности — одна из главных причин ухода опухоли из-под иммунологического контроля. Уменьшение их экспрессии нередко ассоциируется со снижением чувствительности не только к лизису ЦТЛ, но и к естественным киллерам. Ключевая роль ТАР и LMP в распознавании обосновывает целесообразность еще одного несомненно перспективного подхода в общей стратегии иммунотерапии — повышение уровня экспрессии указанных белков различными путями: трансфекцией соответствующих генов, действием цитокинов, усиливающих их экспрессию, в частности IFNy и др. Антигены, рестриктированные молекулами I класса ГКГ, могут быть представлены различными путями. Прямая презентация — деградация цитолитических белков с участием протеосом, транспортом пептидов через мембрану эндоплазматического ретикулума и последующей экспрессией комплекса молекула ГКГ—эпитопы антигена опухоли на поверхность опухолевой клетки. Перекрестная презентация включает внутриклеточный процессинг опухолевых антигенов антигенпрезентирующими клетками. Как известно, прямая презентация, как правило, направлена на представление антигена СD8+Т-лимфоцитов, а перекрестная — СD4+Т-лимфоцитов. При этом показано, что перекрестная презентация необходима и для индукции клеток памяти CD8+, однако остается неясным, способна ли такая презентация влиять на цитотоксичность последних. Для ответа на этот вопрос были проведены опыты с индукцией прямой и перекрестной презентации при использовании мутантных антигенов I класса ГКГ, не способных осуществлять презентацию даже нормальных антигенов этого класса. Результаты исследований показали, что первые индуцируют очень слабую цитотоксичность ЦТЛ, а оптимальная индукция цитотоксичности, но не клеток памяти ЦТЛ, осуществляется при прямой презентации антигена опухолевыми клетками. Как уже указывалось, в структуру антигенов I класса ГКГ наряду с локусами А, В, С входят и другие локусы, в частности G, E и F, которые, в отличие от антигенов локусов А, В, С, характеризуются ограниченным полиморфизмом и поэтому называются неклассическими молекулами. Они отличаются от классических не только ограниченным полиморфизмом, но и особенностями транскрипции, экспрессии и иммунологическими функциями. Антигены локуса G (не принимающие участия в классическом распознавании) экспрессируются трофобластами, на поверхности которых обычно отсутствуют антигены других локусов ГКГ. Физиологическая роль HLA-G в этих случаях заключается в ограничении роста клеток, включая трофобласты, благодаря чему эти антигены играют важную роль в установлении толерантности плода к иммунологической системе матери. Интерес к выявлению антигенов локуса G на опухолевых клетках возник сравнительно недавно и большой вклад в понимание значения экспрессии HLA-G внесли P. Paul и сотрудники. Стало известно, что HLA-G может находиться в мембранно-связанной и растворимой формах, что определяет наличие его различных изоформ: HLA-G1, HLA-G2, HLA-G3, HLA-G4 — изоформы, связанные с мембраной, HLA-G5, HLA-G6, HLA-G7 — растворимые изоформы; некоторые из них обнаруживаются как в супернатантах культивируемых клеток, так и в различных жидкостях организма. Естественно, что сравнительная новизна этого вопроса оставляет неясными многие детали, касающиеся оценки значения экспрессии антигенов локуса G. Тем не менее, несмотря на некоторую неоднозначность такой оценки, полученные результаты позволяют достаточно определенно установить важность экспрессии молекул HLA-G опухолевыми клетками и могут быть использованы для понимания процессов лизиса цитотоксическими клетками. Последнее объясняется, в основном, тем, что взаимодействие с HLA-G приводит к ингибиции лизиса опухолевых клеток, формированию толерантности, что можно рассматривать как благоприятные условия для ухода опухоли из-под иммунологического контроля. Возможный уход от лизиса опухолевых клеток, которые экспрессируют HLA-G, очевидно, связан с ингибицией рецепторов, ответственных за цитотоксичность. В последнее время стали известны несколько типов таких ингибиторных рецепторов, впервые один из них был описан в начале 90-х годов. Более подробные сведения об ингибиторных рецепторах будут изложены ниже. Стало известно, что ингибиторные рецепторы взаимодействуют с молекулами HLA-G и таким образом способствуют уходу опухоли из-под иммунологического контроля. Возможность этого усиливается и тем обстоятельством, что ингибиторные рецепторы экспрессируются на различных циготоксических лимфоцитах: Т-лимфоцитах, естественных киллерах и естественных киллерных Т-лимфоцитах. О неоднозначности трактовок значения экспрессии антигенов HLA-G для процесса распознавания свидетельствуют также результаты изучения значительного количества образцов различных опухолевых тканей и клеток многих опухолевых линий с целью выявления экспрессии антигенов А, В, С, а также G и его изоформы — G1. Результаты этих исследований показали, что в небольшом числе случаев наблюдается транскрипция мРНК антигенов локуса G при отсутствии экспрессии его изоформы — G1. Итогом этих исследований было заключение, что антигены HLA-G, и в частности его изоформа G1, либо не играют роли в осуществлении ингибиторного сигнала киллерных клеток, либо эта роль ничтожно мала. К аналогичным выводам при изучении экспрессии HLA-G клетками меланомы пришли и другие исследователи. Было установлено, что клетки меланомы экспрессировали этот антиген только de novo, что дало основание рассматривать экспрессию локуса HLA-G на клетках меланомы не как закономерную. Эти же исследователи показали, что IFNyHe влияет на экспрессию антигенов HLA-G и поэтому терапия данным цитокином не способствует уходу опухоли от лизиса. Несмотря на то, что авторы указанных исследований не дают окончательной оценки значения экспрессии HLA-G, они не исключают, что экспрессия этих антигенов может препятствовать развитию тех проявлений противоопухолевого иммунитета, которые способствуют опухолевой прогрессии. Такое заключение было сделано при исследовании клеток меланомы, на которых был установлен высокий уровень сплайсинга HLA-G-транскрипции, сочетающийся с прогрессированием опухолевого роста. Молекулы HLA-G могут экспрессироваться на активированных макрофагах и дендритных клетках, инфильтрирующих карциному легкого, а также легочную ткань при других патологических процессах. Предполагается, что экспрессия HLA-G этими клетками может препятствовать презентации антигена и благоприятна для прогрессии как злокачественного роста, так и воспалительных процессов. Некоторые авторы склонны рассматривать экспрессию HLA-G как фактор ускользания опухоли из-под иммунологического контроля даже в тех случаях, когда проведенные исследования не дают прямых доказательств для такого заключения. Например, при исследовании клеток (свежевыделенных и клеток различных линий) гепатомы, меланомы, карцином не выявлено экспрессии антигенов HLA-G. Отмечено также, что опухоли не были инфильтрированы естественными киллерами и лизис опухолевых клеток не наблюдался. Тем не менее авторы не исключают возможной роли антигенов ГКГ локуса HLA-G в процессе ускользания опухоли от иммунологического контроля. Установлено, что HLA-G-молекулы в большем количестве случаев экспрессируются макрофагами и ДК, инфильтрирующими карциному легкого, чем при незлокачественных заболеваниях. По мере изучения роли экспрессии HLA-G сомнения относительно ее значения уменьшались, и в настоящее время есть основания считать, что экспрессия HLA-G может: 1) быть дополнительным механизмом ускользания опухоли от иммунологического контроля; 2) вызывать иммунологическую толерантность; 3) ингибировать цитотоксичность киллерных клеток. Если же учесть, что HLA-G может ингибировать лизис различными киллерными клетками, то спектр возможных негативных влияний экспрессии этих молекул значительно расширяется. К не классическим антигенам системы ГКГ относятся и молекулы локуса E — HLA-E. Эти молекулы характеризуются ограниченным полиморфизмом и с высокой специфичностью связывают пептид Ia, который происходит из полиморфных классических молекул А, В, С и стабилизирует белки ГКГ, способствуя их продвижению к клеточной мембране. Исследование кристаллической структуры HLA-E показало, что он обладает способностью связываться с пептидами Ia HLA-1 при участии белков-транспортеров (TAP-зависимым путем), может взаимодействовать с рецепторами естественных киллеров, ингибируя их лизис. Специфичность связывания молекул локуса E с Ia определяется внутренними свойствами молекулы HLA-E. Подобно молекулам антигена HLA-G, молекулы антигенов HLA-E также выявляются на трофобластах, тормозят активность естественных киллеров и рассматриваются как компонент защиты от распознавания материнскими ЦТЛ; при определенных условиях антигены HLA-E могут активировать естественные киллеры. Если в эндоплазматическом ретикулуме нет основного пептида, то молекулы локуса E теряют стабильность и деградируют еще до достижения поверхности клетки. Если в клетках происходят изменения (в результате попадания инфекции, злокачественной трансформации), снижается экспрессия А, В, С или ингибируется активность ТАР, молекулы локуса E также могут не достигать поверхности. Молекулярные механизмы определения функции антигенов локуса E подлежат дальнейшему изучению. Однако при наличии ряда невыясненных вопросов есть данные о строгой зависимости между экспрессией антигенов локуса E и ко-экспрессией β2m. Как отмечалось, описан еще один локус молекул антигенов I класса ГКГ — локус F. Информация об этом локусе очень ограничена, а сравнительное исследование экспрессии антигенов локуса F у обезьян и человека показало, что он выявляется только у человека. Данных о роли молекул локуса F в распознавании опухолевых антигенов нет. Заканчивая изложение данных о классических и неклассических молекулах, нельзя обойти вниманием и недавно полученные факты, что растворимые формы как классических, так и неклассических молекул, в частности HLA-G, могут индуцировать апоптоз активированных СD8+Т-ли мфоцитов. Изучение этой апоптозиндуцирующей способности в отношении активированных СD8+Т-лимфоцитов показало, что их связывание с растворимыми формами как классических, так и неклассических антигенов приводит к усилению Fas/FasL-взаимодействия, секреции растворимой формы FasL СD8+Т-лимфоцитами, что сопровождается ингибицией цитотоксичности этих клеток. Авторы предполагают, что растворимые формы указанных антигенов выполняют иммунорегуляторную роль в различных условиях, включая и ряд заболеваний, который характеризуется активацией клеток системы иммунитета и повышением уровня sHLA-A, sHLA-B, sHLA-C, sHLA-G в сыворотке крови. Для понимания значения экспрессии антигенов I класса ГКГ важен факт, согласно которому уровень экспрессии антигенов ГКГ по-разному влияет на индукцию цитотоксичности различных киллерных клеток. Так, для оптимального лизиса опухолевых клеток ЦТЛ необходим высокий уровень антигенов I класса ГКГ, в то время как эффективный лизис другими киллерными клетками, в частности естественными киллерами, может осуществляться и при низком уровне указанных антигенов ГКГ, что показано в опытах с аденокарциномой кишечника мышей. Изменения экспрессии антигенов ГКГ (преимущественно снижение) выявлены при многих предопухолевых состояниях, что особенно отчетливо проявляется у антигенов I класса ГКГ. Причины этого снижения могут быть различны: мутации соответствующих генов, контролирующих экспрессию антигенов I класса ГКГ, нарушение регуляции презентации антигенов с участием антигенов I класса ГКГ, ингибиция гликолизирования или транспорта молекул I класса ГКГ, мутации в TAP-генах, мутации или перераспределение в β2m, изменение в структуре хроматина антигенов I класса ГКГ, экспрессия онкогенов и снижение уровня экспрессии молекул ГКГ под влиянием вирусов и др. Достаточное количество данных показывает, что снижение уровня экспрессии антигенов I класса ГКГ часто наблюдается при таких предопухолевых патологиях, как дисплазии, кандиломы, папилломы. Однако это наблюдается не при всех предопухолевых состояниях. Например, при кондиломах, раке шейки матки, молочной железы, гортани и наличии соответствующих генетических и морфологических изменений экспрессия антигенов ГКГ I класса не нарушена. Более того, в некоторых случаях, например, при аденомах кишечника, которые, как известно, характеризуются аккумуляцией таких онкогенов, как k-ras, экспрессия антигенов гистосовместимости не изменена. Наличие экспрессии антигенов I класса ГКГ во многих случаях сочетается с благоприятным прогнозом, например при раке молочной железы, гортани и др. Различные дисплазии, которые сопровождаются снижением экспрессии антигенов I класса ГКГ, в частности с локализацией в шейке матки, органах дыхательного и желудочного тракта, нередко сочетаются с уменьшением экспрессии адгезивных молекул, важных для межклеточных взаимодействий при формировании противоопухолевого иммунитета. Общее представление о динамике экспрессии антигенов I класса ГКГ на нормальных клетках, при предопухолевых состояниях, а также злокачественно трансформированных клетках различных органов дает схема 1.
Различный характер уменьшения экспрессии антигенов отдельных локусов ГКГ I класса демонстрируют исследования, проведенные с клетками инвазивного рака прямой кишки. Исследования показали, во-первых, общую высокую частоту снижения экспрессии этих антигенов (до 40 %) и высокую частоту их повреждения (до 73 %), во-вторых, выявлены локус-специфические различия в повреждении: HLA-A и HLA-B — соответственно в 9 и 8 %, параллельное повреждение HLA-A и HLA-B — в 2 % и не отмечено изменений в экспрессии HLA-C-локуса [73]. Высокую частоту нарушения экспрессии антигенов I класса ГКГ при инвазивном раке прямой кишки авторы рассматривают как благоприятное условие для ускользания опухоли из-под иммунологического контроля. Снижение уровня антигенов I класса ГКГ может быть различным — полным, локусспецифическим или аллелеспецифическим. Обнаружено, что во многих случаях уменьшение экспрессии антигенов I класса связано с формированием резистентности опухолевых клеток к лизису киллерными клетками. Несмотря на то, что снижение уровня экспрессии антигенов I класса ГКГ опухолевыми клетками различного гистогенеза и локализации наблюдается в подавляющем большинстве случаев, возможны и исключения — экспрессия не уменьшается, а в отдельных случаях уровень экспрессии повышается. Тем не менее обращает на себя внимание такой важный факт: в ряде случаев при отсутствии изменений в экспрессии молекул антигенов ГКГ либо даже при ее усилении противоопухолевая иммунологическая защита не формируется. Такая нестандартная ситуация вызывает естественный вопрос: почему при незначительном снижении уровня экспрессии антигенов ГКГ, отсутствии изменений и даже усилении экспрессии противоопухолевый иммунологический ответ все-таки не развивается? Причины этого могут быть различны и будут рассмотрены в последующих разделах. Однако очень важно иметь в виду, что отсутствие формирования противоопухолевого иммунитета еще не означает, что процесс распознавания не произошел. К сожалению, есть убедительные доказательства того, что в некоторых случаях процесс распознавания приводит к индукции другой формы иммунологического ответа — толерантности. При том что, как правило, для представления антигенов опухоли необходимы экспрессия антигенов ГКГ и процессинг опухолевых антигенов имеются наблюдения, согласно которым ослабление процессинга и экспрессии антигенов I класса ГКГ не всегда служит препятствием для лизиса опухолевых клеток соответствующими лимфоцитами. Такие данные получены при исследовании клеток нейробластомы с очень незначительным уровнем экспрессии антигенов I класса ГКГ. Однако даже этого уровня оказалось достаточно для распознавания при условии, что опухолевые клетки были инфицированы вирусом гриппа. Такая чувствительность клеток нейробластомы к действию киллерных лимфоцитов позволяет характеризовать ее как опухоль, чувствительную к иммунотерапии. При всем интересе к этим данным возникают вопросы, на которые сегодня еще нет ответов. Например, можно ли проводить параллель между такими условиями лизиса опухолевых клеток и возможностью лизиса неинфицированных опухолевых клеток со сниженной экспрессией антигенов I класса ГКГ? Можно ли определить минимальный порог экспрессии антигенов ГКГ, который вызывает индукцию иммунологического ответа? При исследовании частоты изменения экспрессии антигенов различных локусов ГКГ I класса показано, что наиболее часто наблюдается уменьшение количества молекул HLA-A, а затем HLA-B; реже имеет место параллельное снижение экспрессии антигенов двух или трех локусов. Обобщая результаты изучения экспрессии антигенов I класса ГКГ с учетом их клинического значения представляется возможным отметить следующее. 1. Существует достоверная отрицательная корреляция между снижением экспрессии антигенов I класса ГКГ и опухолевой прогрессией при многих опухолях — первичной карциноме молочной железы, раке кишечника, шейки матки, ротовой полости и гортани, мочевого пузыря, меланоме. При этой выраженной общей закономерности известны единичные исключения, которые проявляются не только в усилении экспрессии антигенов I класса ГКГ, но даже в появлении этих антигенов на тех клетках, которые до этого их не экспрессировали, что наблюдалось при некоторых опухолях мышечной ткани, в частности при рабдомиосаркоме. 2. Резкое снижение уровня экспрессии антигенов I класса часто совпадает с ранним метастазированием, что особенно характерно для клеток меланомы, у которых, как правило, наблюдается выраженный дефицит экспрессии антигенов I класса ГКГ. 3. Существует корреляция между степенью дифференцировки опухолевых клеток и уровнем экспрессии антигенов I класса ГКГ — степень дифференцировки уменьшается по мере снижения уровня экспрессии. Эти данные подтверждены при параллельном изучении экспрессии различных локусов ГКГ и данных гистологических исследований, которые показали, что наиболее слабая экспрессия антигенов I класса ГКГ сочеталась с низкой дифференцировкой опухолевых клеток, их выраженной инвазивностью и большой метастатической активностью, что особенно отчетливо проявилось при изучении клеток рака носоглотки. 4. Интенсивность снижения экспрессии антигенов I класса ГКГ варьирует в зависимости от локализации опухоли и исходного уровня экспрессии этих антигенов: клетки скелетных мышц и слизистой оболочки желудка могут быть отнесены к клеткам, слабо экспрессирующим антигены I класса ГКГ, а клетки центральной нервной системы практически их не экспрессируют. 5. Нередко снижение уровня экспрессии антигенов ГКГ ассоциируется со слабой иммуногенностью опухолевых клеток. 6. При многих опухолях человека, особенно при меланоме, уровень экспрессии TAP-1 и ТАР-2 уменьшался также LMP, что обусловлено либо их структурными повреждениями, либо дисрегуляцией и ассоциируется с быстрым метастазированием. 7. Снижение уровня экспрессии антигенов ГКГ с полным основанием считают одной из важнейших причин ускользания опухоли из-под иммунологического контроля. 8. Принципиально важна необходимость учета особенностей экспрессии антигенов I класса ГКГ до начала иммунотерапии, что, по мнению многих авторов, может существенно предопределить ее эффективность, в частности вакцинации опухолевыми пептидами.
| |
|
| |
 Уровень экспрессии антигенов ГКГ I класса снижается значительнее по мере развития опухолевого процесса, о чем свидетельствуют многие наблюдения. Нередко уменьшение количества этих антигенов ассоциируется с ускользанием опухоли из-под иммунологического контроля, ранним метастазированием, дессиминацией процесса, что отмечено при меланомах, раке носоглотки, кишечника. Это объясняет, почему во многих случаях наблюдается параллелизм между нарушениями в генах, кодирующих экспрессию антигенов ГКГ, особенностями течения опухолевого процесса и эффективностью иммунотерапии, точкой приложения которой являются Т-лимфоциты. Такое заключение подтверждают наблюдения, согласно которым увеличение частоты нарушения экспрессии антигенов I класса ГКГ может сочетаться либо с отсутствием эффекта иммунотерапии, либо быстрым рецидивированием заболевания. Эти наблюдения свидетельствуют о возможности того, что в основе усиления и распространения опухоли лежит селекция опухолевых клеток, которые приобретают способность ускользать от иммунологического распознавания в связи с нарушениями экспрессии антигенов ГКГ.
Уровень экспрессии антигенов ГКГ I класса снижается значительнее по мере развития опухолевого процесса, о чем свидетельствуют многие наблюдения. Нередко уменьшение количества этих антигенов ассоциируется с ускользанием опухоли из-под иммунологического контроля, ранним метастазированием, дессиминацией процесса, что отмечено при меланомах, раке носоглотки, кишечника. Это объясняет, почему во многих случаях наблюдается параллелизм между нарушениями в генах, кодирующих экспрессию антигенов ГКГ, особенностями течения опухолевого процесса и эффективностью иммунотерапии, точкой приложения которой являются Т-лимфоциты. Такое заключение подтверждают наблюдения, согласно которым увеличение частоты нарушения экспрессии антигенов I класса ГКГ может сочетаться либо с отсутствием эффекта иммунотерапии, либо быстрым рецидивированием заболевания. Эти наблюдения свидетельствуют о возможности того, что в основе усиления и распространения опухоли лежит селекция опухолевых клеток, которые приобретают способность ускользать от иммунологического распознавания в связи с нарушениями экспрессии антигенов ГКГ.| Всего комментариев: 0 | |